Understanding the factors that effect maximal fat oxidation (pdf original)
Purdom T, Kravitz L, Dokladny K y Mermier C
J Int Soc Sports Nutr. 2018 Jan 12;15:3. doi: 10.1186/s12970-018-0207-1
Introducción
Los lípidos son el sustrato principal responsable del aporte de energía durante el ejercicio submáximo. Sin embargo, el rol principal de los lípidos durante la respiración celular aún no está bien definido. El tejido adiposo subcutáneo, los triglicéridos intramusculares (IMTG), el colesterol y los ácidos grasos de la dieta coparticipan en el proceso de oxidación de ácidos grasos (FAox). Además de la energía aportada por los lípidos, en ejercicios submáximos los hidratos de carbono contribuyen en el aporte de energía (CHOox). Sin embargo, a medida que aumenta la intensidad del ejercicio, la contribución de los hidratos de carbono aumenta en relación a la de los lípidos. No obstante, la oxidación de ácidos grasos es el combustible predominante durante el ejercicio submáximo (<65%VO2max). Incrementos en la intensidad del ejercicio por encima del 64% del VO2max provocan un cambio en la contribución energética favoreciendo la CHOox. El término usado para describir el punto de máximo uso de lípidos es máxima oxidación de grasas (MFO). Ejercicios que exceden MFO involucran una mayor proporción de oxidación de CHO. El punto de máxima oxidación de grasas ha sido descrito entre el 47 y 75% del VO2max y varía entre sujetos entrenados y no entrenados y entre mujeres y hombres. No obstante, MFO ha sido establecido en rangos que van desde 0.17-1.27 g/min, donde sujetos adaptados a la cetogenia pueden superar valores de 1.5 g/min. Entre los factores que alteran el ratio de oxidación de grasas se encuentran: el nivel de entrenamiento, la intensidad del ejercicio, la duración del ejercicio, el sexo y el aporte nutricional. Cualquiera de estos factores facilita o inhibe cambios fisiológicos que influyen en la FAox y son discutidos en las siguientes secciones.
Oxidación de los lípidos
Lipólisis
Los triglicéridos son la forma más corta de lípido que podemos encontrar en los adipocitos y el músculo estriado, los cuales están formados por una molécula de glicerol (molécula con 3 carbonos) unida a tres ácidos grasos (FA). Los ácidos grasos son cadenas de carbono unidas entre si acompañados de átomos de hidrógeno. El proceso intracelular que permite liberar los FAs de la columna de glicerol se denomina lipólisis. Una vez que ha tenido lugar este proceso, los FAs son liberados en el torrente sanguíneo y son transportados hacia el músculo activo para su oxidación. Las reservas de tejido adiposo pueden acumular una cantidad significativa de TAG y suministrar un aporte interminable de energía para un rendimiento prolongado durante el ejercicio. Una persona con 7-14% de grasa corporal tiene >30,000 kcal de reserva de energética almacenadas en el tejido adiposo. Por lo tanto, si la intensidad del ejercicio se mantiene por debajo del 65% del VO2max el ejercicio teóricamente puede prolongarse durante un tiempo indefinido debido a la oxidación de las reservas endógenas de TAG. Sin embargo, cuando las intensidades de ejercicio superan 65% VO2max, la FAox se reduce, aumentando la dependencia energética de CHO. El proceso de lipólisis se controla en gran medida a través de sistema endocrino. La liberación de epinefrina estimula la lipólisis y por lo tanto aumenta las concentraciones séricas de FA. En reposo, concentraciones de catecolaminas (epinefrina) en sangre son bajas. A medida que la intensidad del ejercicio aumenta, se produce un aumento simultáneo y progresivo de epinefrina desde las glándulas suprarrenales. Dependiendo de la intensidad y/o duración del ejercicio, las concentraciones de catecolaminas pueden superar 20 veces los niveles basales. El ejercicio provoca la liberación de catecolamina que a su vez estimula la lipólisis, liberando los FA de la molécula de glicerol. Durante intensidades de ejercicio equivalentes al 60% VO2max, las concentraciones de FA séricas superan 2-3 veces los valores de reposo. La unión de epinefrina al receptor β-adrenérgico en las membranas de las células adiposas desencadena una cascada de eventos que comienzan con la fosforilación de la adiposina trigliceril lipasa (ATGL). Hallazgos recientes indican que la lipólisis está bajo una regulación jerárquica por parte de la ATGL y hormonas sensibles a la lipasa (HSL). Además, estudios han demostrado que ATGL tiene una mayor sensibilidad a la epinefrina (un aumento de 10 veces) en comparación con HSL. Por lo tanto, ATGL disocia el primer FA de la molécula de glicerol formando diacilglicerol + FA o (DAG), mientras que HSL es responsable de la segunda disociación de la cadena de FA. Por último, el catabolismo del monoacilglicerol se facilita gracias a la acción de la monoglicerol lipasa, donde el FA es trasportado y el glicerol es utilizado en las vías glucolíticas o gluconeogénicas, principalmente en el hígado. Los FA del músculo esquelético endógeno, denominados IMTGs, pueden contribuir a la FAox global independientemente de la contribución de FA séricos. Los triacilglicéridos intramusculares se disponen dentro del músculo estriado, principalmente en las fibras de tipo I muy próximos a las mitocondrias. El proceso de liberación de FA intramusculares de la molécula de TAG para su oxidación es algo diferente al del tejido adiposo periférico. El transporte a través de la membrana celular no es un limitante en la oxidación de IMTG debido a que se almacenan dentro de la célula muscular. Sin embargo, la lipoproteína lipasa (LPL) y las HSL son necesarias para movilizar los FAs (lipólisis) desde la molécula de glicerol intracelular. La lipoproteínlipasa es una lipoproteína unida al endotelio capilar intramuscular, y es responsable de liberar el primer FA de la molécula TAG dentro de la célula, formando DAG. El proceso de oxidación de IMTG es facilitado por las HSL y es similar al proceso de oxidación del tejido adiposo subcutáneo derivado de HSL. La HSL tiene tres características importantes que afectan a la oxidación de DAG. En primer lugar, HSL demuestra una afinidad 10 veces mayor a DAG en comparación con TAG. En segundo lugar, HSL funciona de forma óptima a un pH de 7,0 y su actividad aumenta a medida que aumenta la intensidad del ejercicio. Por último, la HSL es estimulada directamente por la epinefrina y es independiente de la cascada energética de cAMP conocida por estimular la lipólisis. A pesar de la presencia conocida de IMTG dentro del músculo (principalmente con sujetos entrenados en resistencia y sujetos diabéticos tipo II), la concentración total de IMTG y su contribución energética todavía está bajo debate debido a las variabilidades tisulares. De forma especulativa el ~10% de los FA derivados del suero se utilizan para reponer IMTG durante el ejercicio. Esto hace que sea difícil cuantificar la contribución real de las IMTGs como sustrato. Además, la variación en las metodologías, por ejemplo, biopsia muscular, trazadores isotópicos, resonancia magnética espectroscópica hacen difícil poder establecer comparaciones. Por último, la disparidad en el estado de entrenamiento y la especificidad dietética de macronutrientes complican aún más la obtención de conclusiones definitivas. Se necesita una mayor evidencia en el ámbito del flujo de energía por parte de IMTG para determinar la influencia de IMTG en la contribución energética durante el ejercicio.
Transporte de ácidos grasos
Las limitaciones en la FAox se deben en parte a que se trata de un sistema de entrega multifacético que tiene una serie de eventos regulatorios. Una vez que los FAs dejan el adipocito, primero se unen a la albúmina, que puede unir hasta 12 moléculas de FA. Curiosamente, debido a la mala circulación en el tejido adiposo periférico y una mayor proporción de FA:albúmina tras el ejercicio, la capacidad de unión a la albúmina puede colapsarse y los altos niveles de ácidos grasos libres en el suero plasmático pueden derivar en una estado de riesgo. Debido a la mala circulación en diabéticos tipo II, un alto porcentaje de FA liberados como resultado de la lipólisis estimulada por catecolamina e inducida por el ejercicio no se liberan en la circulación durante el ejercicio de alta intensidad. Sin embargo, se ha demostrado que el entrenamiento de resistencia aumenta el flujo sanguíneo al tejido adiposo subcutáneo en 2 a 3 veces, lo que puede aumentar el transporte general de FA al músculo activo. A pesar del efecto positivo del entrenamiento de resistencia en la circulación de FA, las limitaciones en la tasa de FAox parecen estar mediadas por el transporte celular más que por el transporte sistemático de FA al suero desde el tejido adiposo. El transporte de ácidos grasos a través de la membrana de la célula muscular se produce a través de proteínas transportadoras, principalmente la CD36. La CD36 se encuentra dentro de la membrana plasmática solo 1 minuto después del inicio de la contracción muscular. Schenk y Horowitz (2006) describieron que las mujeres obesas sedentarias que entrenan al >70%HRmáx incrementaron la expresión de CD36 en un 25%. Llevar a cabo ejercicio de resistencia se correlacionó con el aumento en CD36 dentro de las membranas celulares musculares (R2=0,857, P <0,003) con un aumento del 23% en FAox en reposo. Además, la regulación positiva de CD36 es rápida y permanece elevada durante 3 días después del ejercicio. Schenk y Horowitz (2006) demostraron que la plasticidad de los cambios celulares debidos al entrenamiento de resistencia influye positivamente en el FAox en reposo (23%) durante los días posteriores a la conclusión del ejercicio. En humanos se ha demostrado que el sexo afecta a la expresión de CD36 debido a las concentraciones circulantes de estrógenos. Después de 90 minutos pedaleando al 60% del VO2max, el ARNm de CD36 fue un 85% más alto en mujeres que en hombres. Curiosamente, existe una capacidad de captación de FA un 49% mayor debido a las mayores concentraciones de proteína CD36 en mujeres entrenadas en comparación con los hombres entrenados. Kiens y col. (2004) afirmaron que las concentraciones de proteína CD36 son un 49% más altas en mujeres que en hombres, independientemente del nivel de entrenamiento. En resumen, el transporte de FA a través de la membrana celular afecta positivamente a la FAox. El entrenamiento de resistencia aumenta la CD36, mejorando el transporte intracelular para la oxidación de ácidos grasos. Mejoras en el transporte de FA al interior celular para su oxidación permite ahorrar CHO tanto para el ejercicio de alta intensidad como para el ejercicio de larga duración.
Transporte intracelular de FA hasta la mitocondria
Dentro de la célula, se ha demostrado que el tipo y la longitud de la cadena de FA determinan las tasas de oxidación dentro de la mitocondria, en gran parte debido a la especificidad del transportador. Existe una relación inversa entre la longitud de la cadena de carbonos de FA y la oxidación, cuando más larga es la cadena de FA, más lenta es la oxidación. Curiosamente, esta relación inspiró la suplementación de ácidos grasos de cadena corta y media (AGCM) como ayuda ergogénica. Sin embargo, aunque se observaron incrementos significativos en FAox con los AGCM en comparación con ACLC, no se observaron diferencias en el rendimiento. Jeukendrup y Aldred sugieren que este hecho puede deberse al rápido transporte y oxidación de MCFAs independientemente de la carnitina palmitotransferasa. Intuitivamente, esto hecho puede parecer ventajoso, sin embargo, se sospecha que el rápido transporte y la oxidación de los AGCM cortos aumentan la producción de cetonas, lo que se opone al aumento del rendimiento durante el ejercicio. Las cetonas son una fuente de combustible viable, reconocida como una adaptación positiva a la dieta cetogénica, sin embargo, el ejercicio de alta intensidad se basa principalmente en el metabolismo glucolítico para el aporte de ATP y por tanto podría verse comprometido. Este concepto es discutido con más detalle en las siguientes secciones. La oxidación lenta de IMTG provenientes de la sangre y de FA de cadena larga (LCFAs) (<12 carbonos) requiere de una proteína transportadora mitocondrial. La proteína transportadora conocida como carnitina palmitoiltransferasa-1 (CPT-1) se encuentra en la membrana mitocondrial externa y es responsable del transporte de LCFA dentro de las mitocondrias Fig. 1. Los ácidos grasos con 12 carbonos o menos se clasifican como cortos o MCFA y pueden pasar a través de la membrana mitocondrial sin transportador. No obstante, el CPT-1 es necesario para el transporte de LCFA, un producto de la carnitina libre, y se encuentra tanto en el citosol como en la matriz mitocondrial Fig. 1. La concentración de CPT-1, localizada dentro de la membrana mitocondrial durante el ejercicio, parece estar regulada en parte por la intensidad del ejercicio. No se observaron cambios significativos en las concentraciones de CPT-1 en sujetos que realizaban ejercicios con intensidades de ejercicio bajas (50% de VO2max) en comparación con el reposo. Sin embargo, se ha demostrado que el ejercicio al 60% de VO2max aumenta las concentraciones de CPT-1. Con intensidades de ejercicio >75% VO2max, las concentraciones de carnitina libre muscular disminuyen progresivamente y, por lo tanto, CPT-1 puede ser una limitación en el transporte de FA, reduciendo la FAox a intensidades de ejercicio más altas.
Durante el ejercicio de intensidad moderada, CPT-1 cataliza la transferencia de un grupo acilo de un FA (acil-CoA) y su unión a la carnitina libre a través de la membrana mitocondrial externa formando acil-carnitina. Una vez en el espacio intermembrana, la translocasa facilita el transporte de acil-carnitina a través de CPT-II hasta la membrana mitocondrial interna en cuyo punto se libera la carnitina. Este proceso describe el papel de la carnitina en el transporte de FA a través de la membrana mitocondrial a intensidades de ejercicio bajas y moderadas. Durante el ejercicio de alta intensidad, sin embargo, también se producen grandes cantidades de acetil-CoA a través de la glucólisis rápida que ingresan a la matriz mitocondrial y reemplazan la utilización de ácidos grasos. El resultado del abundante acetil-CoA derivado de la vía glicolítica forma acetilcarnitina y monopoliza el transporte de acil-CoA derivado de FA que limita la carnitina libre disponible. Por lo tanto, la carnitina libre se utiliza para amortiguar el exceso de acetil-CoA derivado de la vía glucolítica formando acetilcarnitina, de este modo, la concentración de carnitina es un factor limitante en el transporte / oxidación de FA (Fig. 1). La intensidad del ejercicio tiene una gran influencia sobre las concentraciones de carnitina libre en músculo activo. En comparación con las condiciones de reposo, se ha demostrado que el ejercicio a intensidades superiores al 75% de VO2max reduce las concentraciones de carnitina libre en el músculo activo en un ~80%. La reducción de carnitina libre durante el ejercicio de alta intensidad se debe a la formación de CPT-1, que actúa como reeptor de FA acil-CoA durante el transporte en la membrana mitocondrial y actúa como tampón al exceso de acetil-CoA derivado de la glucólisis. Por lo tanto, a medida que la intensidad del ejercicio aumenta más allá de la intensidad moderada, la carnitina puede ser una limitación en la utilización de FA como sustrato debido a la amortiguación de la acetil carnitina glucolítica. El resultado del rápido aumento en las concentraciones de acetil-carnitina derivadas de la glucólisis a altas intensidades de ejercicio limita directamente el transporte de FA-acetilo a las mitocondrias, lo que reduce el potencial de FAox.

Oxidación de ácidos grasos
La oxidación de ácidos grasos o beta-oxidación (beta-ox) describe el proceso catabólico de eliminación de iones H+ de los FA mientras se produce la acetil-CoA, que se metaboliza aún más en el ciclo de TCA (Ciclo de Krebs). Una de las enzimas clave de beta-ox conocida como β-hidroxiacil-CoA deshidrogenasa (HAD) está directamente involucrada con la FAox en la mitocondria. Se ha demostrado que el entrenamiento aeróbico y las dietas ricas en grasas aumentan la expresión y la actividad de la proteína HAD. La oxidación de ácidos grasos está directamente influenciada por la actividad de HAD, además de con el transporte de FA a través de las membranas celulares y mitocondriales. Mientras que la FAox fluctúa continuamente, el sistema endocrino es el principal responsable de la regulación de la oxidación de lípidos en reposo y durante el ejercicio. Los mecanismos hormonales que estimulan el metabolismo de los lípidos se basan principalmente en catecolaminas, cortisol, hormona del crecimiento, donde la insulina es inhibidora. Debido a que la FAox tiene una tasa máxima, es importante identificar a qué intensidad de ejercicio se produce la MFO para potenciar la quema de grasa, prescribir ejercicio y para establecer recomendaciones dietéticas. Identificar los estímulos que influyen en la oxidación de las grasas es importante para aportar recomendaciones de ejercicio a intensidades que faciliten un potencial óptimo de quema de grasas.
Factores que influyen máxima oxidación de grasas
Nivel de entrenamiento
El mantenimiento de un nivel de entrenamiento alto afecta el potencial de FAox debido al aumento en los IMTG, a los cambios de proteínas celulares / mitocondriales y a la regulación hormonal. Las adaptaciones que se producen debido al entrenamiento de resistencia regular favorecen la capacidad de oxidar la grasa a cargas de trabajo más altas, además de aumentar la MFO. Se ha demostrado que el aumento de la oxidación de grasa mejora con el entrenamiento de resistencia y, por lo tanto, los aumentos en MFO son paralelos a los cambios en el estado de entrenamiento. Bircher y Knechtle (2004) demostraron este concepto al comparar sujetos obesos sedentarios con atletas y encontraron que la MFO estaba altamente correlacionada con la capacidad respiratoria y, por lo tanto, con el estado de entrenamiento. Los sujetos entrenados poseen una mayor capacidad para oxidar la grasa a mayores intensidades de ejercicio y, por lo tanto, demuestra la correlación entre la capacidad respiratoria y la MFO. Sin embargo, se observa una tasa similar en las concentraciones séricas de glicerol en sujetos sedentarios frente a entrenados. Estos resultados entran en conflicto con los resultados de Lanzi y col. (2014) que demostraron que los sujetos obesos tenían una tasa de FA en suero más alta probablemente debido al aumento de la masa total de tejido adiposo (kg). Además, los sujetos sedentarios / obesos tienen un transporte celular reducido y capacidades de oxidación de grasas, por lo que mantienen concentraciones séricas de FA más elevadas. A pesar de la tasa reducida aparición de glicerol para la población entrenada reportada por Lanzie y col., se demostró que las mujeres entrenadas oxidan la grasa al doble de la tasa normal en comparación con la población obesa. El efecto de entrenamiento, y en consecuencia un aumento en la capacidad respiratoria es en parte el resultado de un aumento en la MFO. Scharhag-Rosenberger y col. (2010) llevaron a cabo un estudio prospectivo para demostrar este concepto utilizando sujetos sedentarios que cumplieron o excedieron las recomendaciones mínimas de ejercicio cardiorrespiratorio del ACSM durante un período de 1 año. La tasa máxima de oxidación de grasa aumentó durante los 12 meses de entrenamiento (preentrenamiento 0.26±0.10; post-entrenamiento 0,33±0,12 g/min) y se produjo a una mayor intensidad de ejercicio (preentrenamiento 35±6% VO2max; entrenamiento 50±14% VO2max). La influencia del nivel de entrenamiento en la MFO también se aplica a las poblaciones entrenadas. En sujetos moderadamente entrenados vs altamente entrenados, la intensidad del ejercicio (%VO2max) a la que se produjo el MFO no fue significativamente diferente, pero la MFO fue elevada para los sujetos altamente entrenados (0,29 ± 10 frente a 0,47±0,17 g/min, respectivamente). Además, las enzimas mitocondriales citrato sintasa y HAD se observaron significativamente más elevadas (49% y 35%) en participantes altamente entrenados versus los moderadamente entrenados respectivamente. El aumento de HAD eleva directamente la tasa de beta-oxo mientras que la citrato sintasa aumenta la tasa del ciclo de Krebs. Este hecho sugiere que la lipólisis y el suministro sistémico de FA no son limitaciones para FAox a intensidades de ejercicio más altas. Por lo tanto, las proteínas de transporte celular FA (CD36 y CPT-1) y la densidad mitocondrial (HAD) son probablemente los factores limitantes en la FAox durante el ejercicio de alta intensidad. Elevar el potencial de FAox aumentando la capacidad de respiración celular aumenta la FAox a intensidades de ejercicio más altas, lo que puede tener una influencia positiva en la capacidad aeróbica. Reconociendo la existencia de grandes diferencias interindividuales en MFO, las diferencias en MFO en relación con el nivel de entrenamiento siguen observándose. Lima-Silva y col. (2010) demostraron que las diferencias en el potencial oxidativo de los lípidos pueden aparecer en los corredores de alto nivel en comparación con los moderadamente entrenados. Sin embargo, aunque no se observaron diferencias estadísticas entre los grupos a la intensidad del ejercicio a la que se observó la MFO, hubo una mayor capacidad para oxidar la grasa en los sujetos altamente entrenados. Vale la pena señalar que el aumento en el rendimiento en corredores altamente entrenados probablemente se atribuye a un mayor potencial oxidativo de CHO a mayores intensidades de ejercicio para mantener cargas de trabajo constante más elevadas. Más tarde, la expresión de proteínas celulares, la capacidad de oxidación y en consecuencia, el estado de entrenamiento, tienen la capacidad de influir en la oxidación de grasas. El nivel de entrenamiento influye aún más en la máxima capacidad oxidativa de la grasa al aumentar las concentraciones de sustrato endógeno. El entrenamiento de resistencia mejora las concentraciones de IMTG en las fibras tipo I hasta tres veces más en comparación con las fibras de tipo II. El aumento en la capacidad de MFO debido al entrenamiento de resistencia está más influenciada por la liberación de IMTG por HSL y proteínas LPL, que son responsables de la liberación de FA intramuscular desde la molécula de IMTG. Sin embargo, durante el ejercicio, el grupo de IMTG se repone constantemente con FA derivadas del plasma. No obstante, el aporte de IMTG durante el ejercicio submáximo que duran <2h es esencial para mantener las cargas de trabajo. El efecto en la duración del ejercicio podría deberse a la saturación del receptor β-adrenérgico, que ocurre durante períodos prolongados de ejercicio. Además, se ha demostrado que la actividad de HSL aumenta inicialmente entre los 10-60 minutos, pero vuelve a los niveles de reposo después de 120 minutos de ejercicio, aumentando la dependencia de FA derivadas del plasma. Una mayor evidencia en el área de la dependencia hormonal en la cinética de los FA ha sido establecida como una limitación en la comprensión del efecto del nivel de entrenamiento en la FAox.
Intensidad
Se ha observado que la intensidad del ejercicio a la que se produce la MFO oscila entre el 45-75% del VO2max, un estudio reciente evaluó la tasa de MFO de 1121 atletas de diversas disciplinas (fútbol americano, triatlón, golf, fútbol, deportes de motor, deportes de travesía y deportes acuáticos, entre otros) encontrando variaciones entre el 23 y el 89% del VO2max. Factores tales como el nivel de entrenamiento, el sexo y la nutrición influyen en la cinética de FAox y, por lo tanto, en la intensidad del ejercicio a la que se observa la MFO. La intensidad del ejercicio ejerce la mayor influencia sobre la MFO en base a una combinación de eventos, entre los que se incluyen cambios en el transporte de FA y la fluctuación hormonal, lo que puede provocar un aumento en la tasa lipolítica. Ciertos cambios celulares y hormonales que ocurren durante el ejercicio están directamente relacionados con la intensidad del ejercicio, lo que puede influir en FAox. La oxidación de los ácidos grasos varía según la intensidad del ejercicio, este hecho justifica la evaluación de la intensidad específica a la que se produce la máxima oxidación de ácidos grasos. A un VO2max del 25%, FAox comprende >90% del aporte energético, concretamente, los FA del plasma proporcionan la mayor contribución de energía, donde el glucógeno muscular y los IMTG contribuyen de forma reducida. A intensidades de ejercicio <65%VO2max. el glucógeno muscular y la oxidación de IMTG aumentan considerablemente hasta generar un 50% del aporte energético, dependiendo de la duración del ejercicio. Bergomaster y col. (2008) compararon 6 semanas de entrenamiento de intervalos de alta intensidad (Wingate Tests) con entrenamiento de continuo de resistencia (~65% VO2max), no encontrando diferencias en la MFO. Estos hallazgos sugieren que el entrenamiento ≥65% VO2max no aumentará el potencial de MFO, lo que está en discordancia con estudios más recientes. Estudios más actuales sugieren que el entrenamiento a intensidades de ejercicio altas influye mucho en la utilización del sustrato. Vale la pena señalar que Bergomaster y col. utilizaron sujetos moderadamente entrenados (VO2max = 41,0 ± 2,0 ml/kg/min) donde Achten y col. (2004) y Nordby y col. (2005) utilizaron sujetos más entrenados (VO2max = 58,4±1.8 y 56,6±1.3 respectivamente) para formular sus conclusiones. Incrementos en la expresión de los transportadores de FAox y proteínas oxidativas (CD36, CPT-1, HAD, etc.) conlleva un aumento en la FAox como resultado de la intensidad del ejercicio. Bergomaster y col. (2008) sugieren que es necesario un volumen mínimo de entrenamiento de dos semanas, independientemente del estado de entrenamiento para que se produzca una adaptación celular suficiente. Los datos aportados por Lima-Silva y col. (2010), sin embargo, muestran que en una muestra heterogénea de sujetos altamente vs moderadamente entrenados (VO2max de 68,4 ± 4,5; 58,6 ± 5,4 ml/kg/min respectivamente) durante un periodo mínimo de 3 años a diferentes intensidades, existe una diferencia del 62% en las tasas de oxidación de grasas (0,47±0,17; 0,29± 0,10 g/min, respectivamente). Por lo tanto, el potencial de mejora en la FAox se relaciona con el entrenamiento a intensidades de ejercicio altas en lugar de una adaptación crónica al ejercicio. Además, también se ha demostrado que las concentraciones de carnitina son una limitación directa en la FAox (Fig. 1) a intensidades de ejercicio altas (>65% VO2max), tanto con IMTG como con FA procedentes de la sangre, independientemente de actividad enzimática mitocondrial tanto en sujetos no entrenados como en sujetos moderadamente entrenados. Curiosamente, los esfuerzos para demostrar la aplicación de la carnitina libre en la MFO a altas intensidades de ejercicio han sido infructuosas. La suplementación con carnitina exógena aumentó la carnitina muscular en un 21% después de 4 semanas de suplementación, no observándose diferencias en el rendimiento deportivo. Aunque la FAox no se midió, los aumentos en la carnitina muscular fueron capaces de amortiguar el exceso de acetil CoA al formar acetilcarnitina y así aumentar la actividad de la piruvato deshidrogenasa (PDH) (38%) al 80% de VO2max. La intensidad del ejercicio también puede influir en la MFO al provocar cambios en las concentraciones de catecolaminas que tienen efectos reguladores sobre la lipólisis, la glucogenólisis y la gluconeogénesis. El aumento en las concentraciones de epinefrina que aumentan paralelamente a la intensidad del ejercicio estimula tanto la glucogenólisis como la gluconeogénesis. A medida que aumenta la intensidad del ejercicio también aumentan las concentraciones de catecolaminas, lo que facilita un aumento simultáneo en la CHO y FAox desde la sangre. Sin embargo, el cuerpo favorece la FAox a intensidades de ejercicio <65% VO2max. Cuando la intensidad del ejercicio excede MFO, FAox (g/min) comienza a disminuir; este proceso se describe con el concepto de cruce (Fig 2).
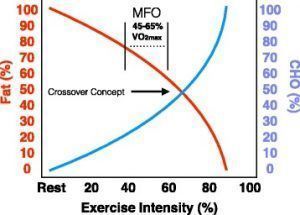
El concepto del punto de cruce representa un medio teórico para comprender el efecto de la intensidad del ejercicio en el equilibrio entre la oxidación de CHO y FA (figura 2). De una forma más específica, el concepto de cruce describe el punto en el que la intensidad del ejercicio requiere una mayor aportación de CHO en detrimento de la FAox. Las limitaciones en la FAox a intensidades altas se deben a la gran cantidad de acetil-CoA producida por la glucólisis rápida. El aumento abrupto en la producción total de acetil-CoA a altas intensidades provocado por la glicólisis suprime el potencial de transporte mitocondrial de FA, reduciendo la FAox (Figura 1). La gran variabilidad intra-sujeto en el punto de cruce a una intensidad de ejercicio dada se puede atribuir en gran parte al nivel de entrenamiento. Se ha demostrado que el nivel de entrenamiento afecta a la liberación de catecolaminas, a la sensibilidad de los receptores, a las concentraciones de sustrato endógeno y a la expresión de proteínas transportadoras; contribuyendo todo ello a generar una variabilidad en la intensidad a la que se produce la MFO. Sin embargo, la MFO aparece en todas las poblaciones, independientemente del nivel de entrenamiento, la nutrición etc., pero está dictada en gran medida por la intensidad del ejercicio.
Duración
Otro factor que influye significativamente en la FAox es la duración del ejercicio. A lo largo de una sesión de ejercicio prolongado, los cambios en las concentraciones de sustrato hormonal y endógeno desencadenan cambios sistemáticos en la oxidación del sustrato. Estudios demuestran que el entrenamiento de resistencia promueve la dependencia de las fuentes de combustible endógenas durante ejercicio de tipo submáximo de una duración de hasta 120 minutos. La duración del ejercicio tiene un gran efecto sobre el origen de los FA con fines oxidativos. Aunque en el inicio del ejercicio la energía depende en gran medida de las fuentes endógenas de combustible (IMTG y glucógeno), se ha demostrado que las reducciones en las concentraciones de IMTG ocurren cuando la duración del ejercicio excede los 90 min. Más allá de los 90 min de ejercicio submáximo (≥65% de VO2max), la oxidación de IMTG se mitiga por el aumento en las LCFA derivadas del suero. Se observaron aumentos tanto en las concentraciones de epinefrina como de LCFA en plasma cuando el ejercicio excedió los 90 min con una reducción simultánea de la actividad HSL. Por lo tanto, el aumento en sangre de LCFAs y la saturación de HSL por la epinefrina se postulan como inhibidores de la HSL reduciendo la oxidación de IMTG cuando el ejercicio excede los 90 min. Cuando el ejercicio excedió los 120 min, la oxidación de IMTG volvió a los valores de reposo y fue contrarrestada por un aumento del 46% en el suministro sérico de FA. Varios estudios muestran que después de 12 h de ejercicio prolongado, las reservas de IMTG se mantienen entre un 50-80% con relación a las concentraciones previas al ejercicio a pesar de la duración extrema del ejercicio. El cambio en las fuentes de combustible de FA intramusculares a FA derivados de la sangre tras 2h de ejercicio submáximo se correlaciona con cambios en las concentraciones de glucosa en sangre. Los sujetos no entrenados que completaron 3 horas de extensiones de rodilla al 60% de 1RM mostraron un aumento del 66% en las concentraciones séricas de glucosa entre la segunda a la tercera hora de ejercicio. Sin embargo, los sujetos entrenados experimentaron una reducción en la captación muscular de CHO durante el mismo marco de tiempo en comparación con los no entrenados. Esto sugiere que los sujetos entrenados fueron capaces de mantener FAox (a pesar del origen del sustrato) durante el ejercicio prolongado para evitar el uso de CHO. Aunque la intervención de ejercicio utilizada en este estudio no se clasifica típicamente como ejercicio de resistencia, el protocolo de ejercicio desmuestra la variación en el origen de la oxidación del sustrato a lo largo del tiempo y amplía los diversos efectos que la duración del ejercicio tiene sobre la oxidación del sustrato. La duración del entrenamiento tiene una gran influencia en la oxidación de FA y CHO durante el ejercicio submáximo prolongado. Sin embargo, el estado de entrenamiento tiene poca influencia en el origen de los FA durante los primeros 120 min de ejercicio submáximo. No obstante, los sujetos entrenados pueden mantener cargas de trabajo más altas con una disminución del trabajo metabólico (FC) durante periodos más largos en comparación con los individuos no entrenados con relación a la capacidad de mantener FAox durante períodos más largos. Además del efecto del nivel de condición física en la FAox, la duración del ejercicio determinará el origen del sustrato durante el ejercicio submáximo.
Sexo
La variabilidad en la FAox debido al sexo se explica debido a las diferencias hormonales inherentes entre hombres y mujeres. En un estudio exhaustivo con más de 300 hombres y mujeres premenopáusicas, la contribución energética de la grasa fue significativamente más alta en las mujeres que en los hombres en todas las intensidades de ejercicio medidas desde 41-61% de VO2max. Los estudios han demostrado consistentemente que las mujeres premenopáusicas tienen una capacidad significativamente mayor para oxidar la grasa durante el ejercicio. Las diferencias entre sexos en la oxidación de grasas durante el ejercicio se atribuyen a la mayor circulación de estrógenos. La evidencia sugiere que el estrógeno estimula directamente la actividad de AMPK y PGC-1α, que se cree que aumentan las proteínas transportadoras de la FAox CD36 y la beta-oxidativa HAD. Además, se ha demostrado que las proteínas beta-oxidativas que participan en la oxidación de LCFA están reguladas en parte por los estrógenos. El resultado del aumento de las proteínas beta-oxidativas está directamente relacionado con un mayor potencial de FAox. Curiosamente, cuando los hombres fueron suplementados con estrógeno, se observaron aumentos en FAox junto con una mayor expresión celular de proteínas beta-ox dentro de los ocho días de la suplementación. Los estrógenos circulantes son naturalmente más altos en las mujeres premenopáusicas en comparación con los hombres. La fluctuación en los niveles de estrógenos es inherente durante todo el ciclo menstrual. Los estrógenos son generalmente más altos durante la fase folicular del ciclo menstrual en comparación con la fase lútea. Paradójicamente, los estrógenos elevados durante la fase folicular no afectan a FAox en comparación con la fase lútea. Sin embargo, los aumentos de estrógenos circulantes endógenos inherentes a las mujeres premenopáusicas aumentan la expresión de proteínas celulares responsables del transporte y la oxidación de FA en comparación con los hombres.
Nutrición
La expresión proteica y la correspondiente oxidación del sustrato endógeno frente a la sistemática varían en función de la ingesta de macronutrientes en la dieta. Se ha demostrado recientemente que las dietas ricas en grasas promueven FAox y tienen capacidades de mejora del rendimiento. Sin embargo, las aplicaciones con relación al tipo de macronutrientes ingerido previo al ejercicio y las mejoras en el rendimiento dependen de una actividad determinada por una duración e intensidad específica. Las dietas que tienen mayores proporciones de un macronutriente específico (por ejemplo, grasa / CHO) han demostrado una mayor capacidad para oxidar el macronutriente primario consumido. Además, las concentraciones endógenas de sustrato aumentan después de la aclimatación a dietas altas en grasas o altas en CHO. Las dietas ricas en grasas aumentan las concentraciones de IMTG al tiempo que disminuyen los niveles de glucógeno dentro del músculo. Alternativamente, las dietas ricas con CHO aumentan las concentraciones de glucógeno, mientras que las IMTG disminuyen. Tras la aclimatación, durante el ejercicio el cuerpo favorece la oxidación de sustratos específicos basado en la adaptación celular a largo plazo (> 48 h). Sin embargo, se ha demostrado que el consumo de macronutrientes predominante después del ejercicio (CHO) influye en la expresión de proteínas celulares en tan solo 2 horas. Cambios crónicos derivados de la expresión celular están influenciados por el tipo de macronutriente ingerido post-ejercicio. Se ha demostrado que la proporción y el periorización en la ingesta de macronutrientes tienen efectos sobre la adaptación celular así como sobre la respuesta fisiológica al ejercicio. Las dietas altas en grasas aumentan el potencial de beta-ox en reposo y durante el ejercicio; sin embargo, las limitaciones de las dietas ricas en grasas (incluida la adaptación a corto plazo (5 dias)) aparecen con el ejercicio de alta intensidad. El ejercicio de alta intensidad (> 75% VO2max) eclipsa el potencial oxidativo de FAox basándose en la glucólisis rápida, o más específicamente, PDH para producir acetil-CoA derivado de la CHO para la síntesis de ATP. La piruvato deshidrogenasa es la enzima responsable de la oxidación del piruvato como sustrato final de la ruta glucolítica. Se ha encontrado la adaptación celular negativa en la actividad de PDH debido a las dietas ricas en grasas, la cual compromete el potencial de rendimiento durante el ejercicio de alta intensidad. Las dietas ricas en grasas (>68% de la ingesta diaria total de calorías) han tenido efectos positivos sobre la reducción de los valores de RER durante el ejercicio de intensidad moderada (~64-70% VO2max) y durante ejercicio prolongado (~3 horas) que indica un aumento en la FAox. La adaptación del organismo a las dietas ricas en grasas permite al cuerpo aumentar el almacenamiento de IMTG y aumentar la FAox. Por el contrario, se demostró que la actividad de PDH y, por lo tanto, la oxidación de CHO se veía comprometida junto con la potencia energética a intensidades de ejercicio ≥70% VO2max. Sin embargo, las aplicaciones de dieta cruzada donde el cuerpo se adaptó a una dieta alta en grasas antes de la carga CHO a corto plazo (36-72 h) mantuvieron las reservas de IMTG mientras que aumentan las reservas de glucógeno, restablecen parcialmente las enzimas glucolíticas, así como restauran parcialmente la CHOox. El aumento en la MFO (g/min) y la intensidad de ejercicio a la que se produce la MFO (% VO2max) es ideal para la mejora del rendimiento durante actividades de larga duración. Sin embargo, una mayor evidencia científica es necesaria para clarificar las reducciones en la actividad celular de PDH tras la adaptación agua a las dietas ricas en grasa y sobre la capacidad de restaurar el potencial glucolítico después de la adaptación a la CHO a corto plazo en ejercicios de alta intensidad intermitente (≥70% VO2max). La alternancia específica de macronutrientes pre-ejercicio tiene el potencial de ser eficaz en la preparación hacia el ejercicio continuado de alta intensidad gracias a la expresión proteica y al almacenamiento adecuado de IMTG y glucógeno muscular. Se ha demostrado que la adaptación a dietas altas en grasa (>50% de grasa total de K/cal) reduce la actividad de PDH en un 59% en reposo y un 29% durante el ejercicio de intensidad moderada (70% VO2max). La reducción en la actividad de PDH debido a las dietas ricas en grasas es un factor limitante en la oxidación de CHO necesario en el ejercicio de alta intensidad a pesar de las reservas energéticas. Sin embargo, cinco días de adaptación a una dieta rica en grasa (~67% de la ingesta total de energía) con un período de carga de CHO a corto plazo de 24 horas (en comparación con una dieta alta en CHO ~70% de ingesta total de energía) mantienen las concentraciones de IMTG y restauran parcialmente la actividad de PDH (71% de una dieta alta en CHO) mientras se mantiene una actividad del 80% en la HSL. No se observaron diferencias en el rendimiento durante una prueba contrarreloj (~10 min al 90%VO2max) entre los sujetos que siguieron una dieta rica en CHO y los que siguieron una dieta alta en grasas con una carga de CHO a corto plazo. Mantener la capacidad de almacenar y oxidar grasa tras la ingesta de una dieta alta en grasas mientras se mantiene la capacidad de oxidar CHO con una carga de CHO a corto plazo proporciona un estado fisiológico ideal para el rendimiento durante el ejercicio de resistencia. Además, la glucogenólisis aumenta durante el ejercicio después de una carga de CHO, lo que indica un aumento en el almacenamiento de glucógeno y una mayor capacidad para producir/mantener la disponibilidad de CHO durante el ejercicio intenso. Investigaciones recientes afirman que las dietas ricas en grasas favorecen la FAox tanto en reposo como durante el ejercicio. Sin embargo, la intensidad del ejercicio dicta la utilización del sustrato, independientemente de la influencia de la dieta, el estado del entrenamiento y la duración del ejercicio. Gracias a este hecho, las dietas altas en grasas cobran sentido durante el entrenamiento de pretemporada cuando los volúmenes de entrenamiento son altos y las intensidades de ejercicio varían entre bajas y moderadas. Sin embargo, durante el ejercicio sostenido de alta intensidad (>70% VO2max) que es común durante la competición, CHO son el sustrato principal en el que se basa el aporte energético. Se necesita una mayor evidencia científica para clarificar el efecto de la manipulación a corto plazo de los macronutrientes sobre las concentraciones endógenas de sustrato, la plasticidad en la expresión celular y la oxidación preferente de un sustrato para determinar si existe un beneficio real sobre el rendimiento durante el ejercicio.
Conclusiones
En resumen, la FAox depende de muchos factores que pueden modificar la expresión celular en un corto período de tiempo. La disponibilidad de macronutrientes, el estado de entrenamiento, el sexo, la intensidad del ejercicio y la duración influyen en la adaptación celular, el transporte sistemático de FA y en la FAox. La intensidad del ejercicio limita la oxidación del sustrato de forma aguda, independientemente del estado de entrenamiento y/o la ingesta nutricional. Además, se necesita una mayor evidencia sobre el momento de ingesta idóneo y el contenido de la ingesta para clarificar qué sustrato podría influenciar favorablemente las adaptaciones fisiológicas en la FAox durante el ejercicio de resistencia. No obstante, las prescripciones de ejercicio y las recomendaciones dietéticas deben tener en cuenta las características específicas de la actividad (duración, intensidad, deporte) para facilitar un plan de entrenamiento que promoviera las adaptaciones idóneas hacia la oxidación del sustrato más relevante en esa prueba que conlleve a su vez una mejora en el rendimiento deportivo.